Долговременная память аплизии
Долговременная память аплизии
Если открытые Кэнделом механизмы процессов привыкания и сенситизации могут служить моделью кратковременной памяти, то какое отношение они могут иметь к долговременной памяти? Группе, работавшей с аплизией, нужно было найти у их излюбленного объекта такой процесс, который определенно можно было бы назвать долговременной памятью и попытаться изучить наподобие того, как это было с успехом сделано в случае привыкания и сенситизации. Этим объясняется повышенное внимание, которое в начале 80-х годов уделялось условнорефлек-торному втягиванию сифона и жабры. Животное обучали реагировать на слабый стимул, обычно не вызывающий этой реакции, таким же образом, как на сильный, который ее уже вызывает (например, на резкое раздражение хвостовой области).
Экспериментальный подход был усовершенствован в 1983 году сотрудником Кэндела Томом Кэрью [8], которому удалось смоделировать аналогичную ситуацию на "упрощенном" препарате. Безусловный раздражитель был заменен повторной активацией сенсорного нейрона, а условный воздействием на клетку серотонина.
Ключевым признаком ассоциативного научения (в отличие от привыкания и сенситизации) служит его долговременный характер, тогда как все рассмотренные до сих пор эффекты были кратковременными. Согласно моему второму критерию, при ассоциативном научении у аплизии должны возникать более стойкие клеточные изменения, соответствующие длительной модификации поведения. Хотя еще в начале 70-х годов было
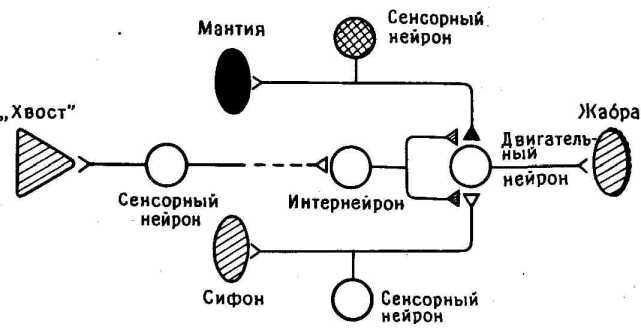
9.3. Схема нервного аппарата классического обусловливания у аплазии. (Один кружок может представлять группу нейронов.) Безусловный раздражитель (резкое воздействие на хвостовую область) возбуждает облегчающие интернейроны, имеющие выходные синапсы на окончаниях аксонов двух других путей: от мантии и от сифона Одновременное слабое воздействие на мантию (условный раздражитель) и сильное на хвостовую область усиливает передачу сигнала от мантии, вследствие чего даже слабое раздражение последней вызывает теперь втягивание жабры; слабое раздражение сифона остается неусиленным и не вызывает реакции втягивания.
показано, что ингибиторы белкового синтеза не влияют на привыкание и сенситизацию, Кэндел только в середине 80тХ годов переключил свое внимание на более долговременные клеточные процессы. Хотя введение этих ингибиторов не сказывается на привыкании, они вызывают амнезию в опытах с ассоциативным научением. Поэтому последнее нельзя осуществить путем кратковременного изменения секреции медиатора; для этого необходим синтез новых белков, и исследователям нужно было идентифицировать их и выяснить их функции в клетке. В связи этим Кэндел предпринял эксперименты вроде тех, которые уже пытались провести исследователи биохимии памяти.
Биохимики добавляли в культуру изолированных ганглиев или клеток радиоактивные предшественники белка и старались выявить образующиеся белковые продукты; те из них, синтез которых начинался или усиливался только при формировании следов памяти, отделяли от множества неспецифических белков. Одновременно обсуждался вопрос, каким образом кратковременные изменения в секреции таких нейромедиаторов, как серотонин, или в транспорте кальция через синаптическую мембрану могут запускать специфический синтез новых белков, необходимых для долговременного запоминания [9].
Клеточные алфавиты или нервная система?
Поскольку все эти проблемы долговременной памяти у моллюсков смыкаются с биохимической проблематикой моих собственных исследований на цыплятах, я намерен на время отложить их обсуждение. Сначала нужно будет заняться теми вопросами, которые с редукционистской прямолинейностью игнорировались в ортодоксальных работах на аплизии (во всяком случае подход здесь был ортодоксальным в середине 80-х годов, хотя сейчас, похоже, он становится все более гибким). Хотелось бы, чтобы мои критические замечания были поняты правильно. В последние двадцать лет группа Кэндела внесла большой теоретический и экспериментальный вклад в изучение клеточных механизмов памяти, но интеллектуальная самоуверенность коллектива в целом и авторитет его руководителя стали приводить к замалчиванию некоторых аспектов всей этой работы и оттеснению тех, кто пытался привлечь к ним внимание. Я не собираюсь переходить на личности и вдаваться в вопросы приоритета, тем более что Сьюзан Олпорт уже вынесла кое-что на публичное обсуждение в книге "Исследователи черного ящика".
Вместо этого я попытаюсь сосредоточиться на некоторых спорных теоретических вопросах.
Кое-кто из недовольных амбициозностью кэнделовской метафоры "клеточного алфавита" ссылается на якобы разную природу научения у беспозвоночных (аплизии) и у позвоночных. Нервная система аплизии состоит из относительно небольшого числа нейронов, однако среди них есть очень крупные. Этим она сильно отличается от мозга позвоночных, который содержит множество мелких нейронов с чрезвычайно большим количеством связей между ними.
Поэтому одно из главных различий в устройстве мозга позвоночных и беспозвоночных иногда видят в том, что у последних на отдельный нейрон или даже синапс ложится гораздо больше работы и ответственности, чем у позвоночных, у которых они распределены между разными клетками. Возможно, так оно и есть, но в последние годы нейрофизиологи-беспозвоночники (те, что изучают беспозвоночных, а не особая группа исследователей, лишенных спинного хребта!), обычно утверждавшие, что излюбленные ими организмы имеют простую нервную систему, несколько изменили формулировку: они говорят теперь о "простых" нервных системах, намеренно употребляя кавычки в знак признания
Морски* улитки и гиппокамп: идеальные объекты? 255
того, что эти системы все же на много порядков сложнее, чем действительно простые компьютерные схемы. У многих беспозвоночных, например у насекомых, нервная система набита крошечными нейронами. Это относится и к таким моллюскам, как осьминоги и кальмары с их крупным мозгом.
Аплизия представляет случай особого рода, ибо это животное легко изучать, но, очевидно, можно без большой натяжки утверждать, что фундаментальные механизмы научения не слишком отличаются у нее от свойственных другим беспозвоночным животным и даже позвоночным с сопоставимыми размерами нервной системы. Синаптические взаимодействия даже 20 000 нейронов достаточно многообразны, чтобы они работали как целостная система, а не отдельные клетки, и некогда популярные утверждения, что в нервной системе насекомых и ракообразных можно найти ключевые "командные" нейроны, оказались столь же несостоятельными, как и пропагандировавшиеся с равным энтузиазмом "командные экономические методы" в Восточной Европе. Иными словами, этот подход так же мало пригоден для организации индивидуального поведения, как и для управления страной.
Другое направление критики представляют некоторые психологи, концентрирующие огонь на вопросе о том, насколько экспериментальная процедура ассоциативного обучения аплизий "действительно" отвечает требованиям классического обусловливания [10]. Однако все эти перебранки по поводу терминологии сейчас интересуют меня гораздо меньше других вопросов, которые я здесь рассмотрю, исходя из критериев необходимости, достаточности и специфичности. Несмотря на поразительную аналогию между процессами привыкания и сенситизации у интактных аплизий и реакциями изолированного сенсорно-моторного синапса аналогию, вполне удовлетворяющую некоторым из моих критериев, здесь имеется явный логический пробел.
Хотя изменения на уровне этого синапса в процессе привыкания при рефлекторном втягивании жабры и сифона действительно происходили, до сих пор формально не доказаны их необходимость и достаточность. Говоря об упрощениях, производившихся группой Кэндела, я уже отмечал, что они устраняли ряд "неудобных" в экспериментальном и теоретическом плане процессов, сопутствующих поведенческой реакции (например, влияние периферической нервной системы и некоторые полисинаптические воздействия на моторные нейроны). Возможны ли привыкание, сенситизация
и ассоциативное научение в случае повреждения ключевых сенсорно-моторных синапсов (пятый критерий)? И только ли эти специфические синапсы изменяются при процессах кратковременного и долговременного научения?
Один из самых упорных критиков Кэндела, нейрофизиолог Кэн Луковяк из Калгари, указывает, что причинная связь между нервной и поведенческой реакциями, постулированная Кэнделом, никогда не подвергалась прямой проверке на интактных животных. Так, например, если сила реакции целиком зависит от отдельного синапса, у интактного животного должна выявляться высокая положительная корреляция между степенью активации специфического моторного нейрона и выраженностью рефлекса втягивания. Попытка обнаружить такую корреляцию Луковяку не удалась.
Создавалось впечатление, что силу рефлекса контролирует не какая-то отдельная клетка абдоминального ганглия, а весь комплекс взаимодействующих клеток как система [11].
Подтверждение этого вывода пришло не от оппонентов, а от самих членов группы Кэндела.
- Долговременная потенциация
- Гиппокамп как когнитивная карта
- Биохимический механизм ДВП
- Можно ли считать двп памятью?
- Никто кроме нас, цыплят
